


Maria Laura Gennaro, M.D. M.Sc.
Professor of Medicine and Epidemiology
marila.gennaro@rutgers.edu | View CV
+1-973-854-3210
room W250 Q-W




NEWS:
- Our work on COVID-19 in children is funded by an R61/R33 award by the NICHD and by a Rutgers subaward within the pediatric RECOVER initiative to study “Long COVID” (OTA 21-015 from the NHLBI).
- Our longitudinal studies on the natural history of SARS-CoV-2 infection have been funded by an R01 award from the NIAID.
- Our research on convalescent plasma therapy for COVID-19 has been funded by an NHLBI emergency administrative supplement.
- Our research on foam cells as therapeutic targets in tuberculosis has been funded by an R01 award from the NHLBI.
- Our FISH-Flow protocol can be found here.
- Our COVID-19 ELISA protocol can be found here.
Mycobacterium tuberculosis is an intracellular pathogen that infects the host macrophage. The most common infection outcome is a standoff in which both pathogen and macrophage undergo adaptive modifications, and survive. Maintaining this equilibrium between bacteria and innate immune cells requires a third player, the host acquired immune (T-cell-mediated) responses. When the T cell response is compromised, the host-pathogen equilibrium is broken, and disease ensues. To date, two billion people carry latent (asymptomatic) M. tuberculosis infection; of these, 10 million become sick every year with tuberculosis, and one and a half million die.
Our laboratory studies mechanisms of adaptation expressed by the bacterium and by the host macrophage during infection, with the goal of finding targets for therapeutic intervention. We also develop tools that may help identify latently infected individuals that are at high risk of developing active tuberculosis. Our expertise in antibody responses to tuberculosis has guided our current effort in studying the antibody response to SARS-CoV-2 and the seroepidemiology of COVID-19.
Macrophage lipid metabolism:
Foam cells are lipid-laden macrophages associated with chronic inflammatory states and tissue damage in many pathological (infectious and non-infectious) conditions. In tuberculosis, foam cells are central to establishing and maintaining chronic Mycobacterium tuberculosis infection. During active tuberculosis, the abundance of foam cells correlates with the extent of tissue damage, linking these cells to immunopathology. Our laboratory studies the biogenesis and key characteristics of foam cells during M. tuberculosis infection, and the interplay between metabolic and inflammatory stimuli leading to their formation.
Statins, which are inhibitors of the mevalonate pathway and de novo cholesterol synthesis, exhibit anti-mycobacterial activity in macrophages infected in vitro and increase the bactericidal efficacy of standard anti-tuberculosis chemotherapy in mice. We are currently studying the mechanisms underlying the observed anti-mycobacterial activity of statins and the interplay between metabolic and inflammatory effects of statin treatment on M. tuberculosis infection. Our work has led to an ongoing, NIH-funded clinical trial in South Africa to assess the effectiveness of statins as adjunctive, host-directed therapy against tuberculosis.
M. tuberculosis stress responses:
Our laboratory also investigates links between regulatory and metabolic adaptive responses of M. tuberculosis during infection. We have shown that expression of lung immunity in mice is associated with upregulation of a “dormancy” regulon, changes in expression of genes involved in central carbon metabolism and respiration, and mycobacterial cell wall remodeling. By collaborating with mathematical modelers and network analysts, we reconstructed the complete sigma factor network of tubercle bacilli and dissected regulatory links and dynamics between the envelope-stress-response network and regulation of central metabolism genes of M. tuberculosis. We also discovered that M. tuberculosis possesses the phage shock protein (Psp) system, a multi-protein system found across bacterial phyla that stabilizes the cell envelope during stress conditions. We study the correlations between Psp function, proton motive force preservation, ATP homeostasis, susceptibility to anti-tuberculosis antibiotics, and mycobacterial virulence. In addition, we are studying the phylogenetic spread, domain architecture, and genomic context of the elements of the Psp system in the bacterial kingdom with the goal of linking evolution of protein and operon structure to function.
FISH-Flow:
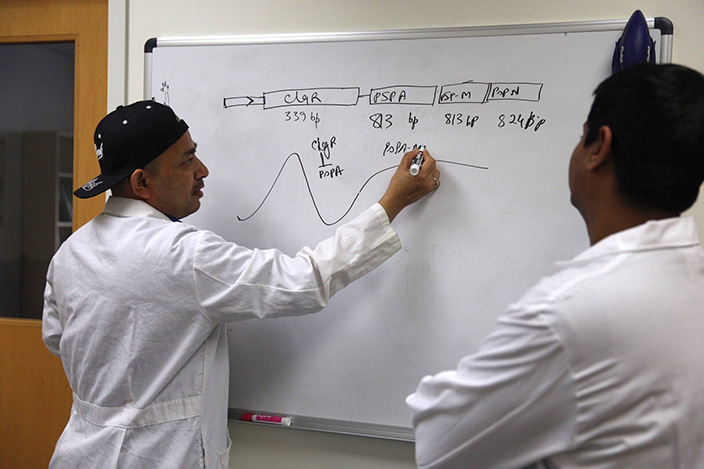
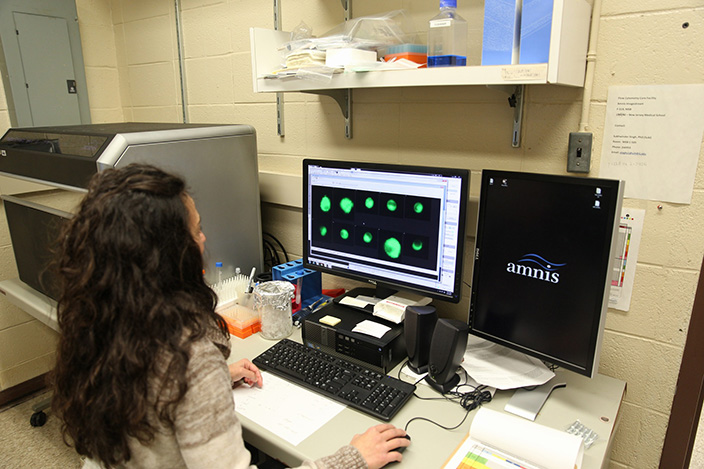
We have developed an RNA-flow cytometry assay (FISH-Flow) which allows to simultaneously measure mRNA and protein in single cells. The method has advantages over in-situ fluorescence microscopy (FISH) and conventional (antibody-based) flow cytometry. Moreover, the nucleic acid probes we use (“Stellaris”) afford extremely low background signals, which makes our protocol more suitable for detection of rare events than RNA flow cytometry techniques utilizing other types of nuclei acid probes. We apply FISH-Flow to detection of antigen-specific T cell responses in tuberculosis. The methodology can also be applied to other cell types, to immune responses in other pathological conditions, and to viral detection. In collaboration with BD Biosciences, we have developed a semi-automated FISH-Flow protocol that reduces labor and cell loss and increases assay reproducibility and its clinical applicability.
COVID-19:
At the beginning of the COVID-19 pandemic, we established a Spike-protein-based ELISA test to detect anti-SARS-CoV-2 antibodies. We have participated in a leadership role in establishing a longitudinal healthcare worker cohort and conducting the serological component of the study. We are also conducting seroprevalence studies to assess the burden of SARS-CoV-2 infection in Essex county, New Jersey. In addition, we are collaborating with basic scientists, pulmonologists, critical care physicians, pediatricians, epidemiologists, hematologists, and clinical trialists across two medical schools and two teaching hospitals to address questions on the relationship between antibody response, memory B cell response, evolution of infection, transmission dynamics, pathogenesis, and clinical manifestations of COVID-19 in adults and children.
Henrich TJ, Montgomery CP, Graf J, Ismali N, Mohandas S, Suthar MS, Brim H, Coffin JM, Pagaria A, Guzmán Rivera J, Vudali U, Keim P, Zhong G, McGrath R, Edwards B, García-Sastre A, Gennaro ML (2025) The role of co-infection in the pathogenesis of acute SARS-CoV-2 infection and development of post-acute sequelae: A perspective. eLife 14: e106308.
Khan R, Ji W, Guzman Rivera J, Madhvi A, Andrews T, Richlin B, Suarez C, Gaur S, Hasan UN, Cuddy W, Singh AR, Bukulmez H, Kaelber D, Kimura Y, Ganapathi U, Michailidis IE, Ukey R, Moroso-Fela S, Kuster JK, Casseus M, Roy J, Burns JC, Kleinman LC, Horton DB, Lakhani SA, Gennaro ML (2025) A genetically modulated Toll-like receptor-tolerant phenotype in peripheral blood cells of children with multisystem inflammatory syndrome. The Journal of Immunology 214: 373–383.
Guzman Rivera J, Zheng H, Richlin B, Suarez C, Gaur S, Ricciardi E, Hasan UN, Cuddy W, Singh AR, Bukulmez H, Kaelber DC, Kimura Y, Brady PW, Wahezi D, Rothschild E, Lakhani SA, Herbst KW, Hogan AH, Salazar JC, Moroso-Fela S, Roy J, Kleinman LC, Horton DB, Moore DF, Gennaro ML (2025) Mass spectrometry combined with machine learning identifies novel protein signatures as demonstrated with multisystem inflammatory syndrome in children. Sci Rep 15: 36843. PMI: 41125642
Guerrini V, Prideaux B, Khan R, Subbian S, Wang Y, Sadimin E, Pawar S, Ukey R, Singer EA, Xue C, Gennaro ML (2025) Cryptococcosis, tuberculosis, and a kidney cancer fail to fit the atherosclerosis paradigm for foam cell lipid content. J Immunol 214: 1358–1369. PMI: 40156376
Bruiners N, Guerrini V, Ukey R, Dikdan RJ, Yang JH, Mishra PK, Onyuka A, Handler D, Vieth J, Carayannopoulos M, Guo S, Pollen M, Pinter A, Tyagi S, Feingold D, Philipp C, Libutti SK, Gennaro ML (2022) Longitudinal Analysis of Biologic Correlates of COVID-19 Resolution: Case Report. Front Med (Lausanne) 9: 915367. PMI: 35783607
Ukey R, Bruiners N, Mishra H, Mishra PK, McCloskey D, Onyuka A, Chen F, Pinter A, Weiskopf D, Sette A, Roy J, Gaur S, Gennaro ML (2022) Dichotomy between the humoral and cellular responses elicited by mRNA and adenoviral vector vaccines against SARS-CoV-2. BMC Med 20: 32. PMI: 35073931
Mishra PK, Bruiners N, Ukey R, Datta P, Onyuka A, Handler D, Hussain S, Honnen W, Singh S, Guerrini V, Yin Y, Dewald H, Choudhary A, Horton DB, Barrett ES, Roy J, Weiss SH, Fitzgerald-Bocarsly P, Blaser MJ, Carson JL, Panettieri RA, Jr., Lardizabal A, Chang TL, Pinter A, Gennaro ML (2021) Vaccination boosts protective responses and counters SARS-CoV-2-induced pathogenic memory B cells. medRxiv. PMI: 33880486
Horton DB, Barrett ES, Roy J, Gennaro ML, Andrews T, Greenberg P, Bruiners N, Datta P, Ukey R, Velusamy SK, Fine D, Honnen WJ, Yin YS, Pinter A, Brooks A, Tischfield J, Hussain S, Jagpal S, Swaminathan S, Parmar V, Reilly N, Gaur S, Panettieri RA, Carson JL, Blaser MJ (2021) Determinants and Dynamics of SARS-CoV-2 Infection in a Diverse Population: 6-Month Evaluation of a Prospective Cohort Study. J Infect Dis 224: 1345-1356. PMI: 34387310
Datta P, Ukey R, Bruiners N, Honnen W, Carayannopoulos MO, Reichman C, Choudhary A, Onyuka A, Handler D, Guerrini V, Mishra PK, Dewald HK, Lardizabal A, Lederer L, Leiser AL, Hussain S, Jagpal SK, Radbel J, Bhowmick T, Horton DB, Barrett ES, Xie YL, Fitzgerald-Bocarsly P, Weiss SH, Woortman M, Parmar H, Roy J, Dominguez-Bello MG, Blaser MJ, Carson JL, Panettieri RA, Jr., Libutti SK, Raymond HF, Pinter A, Gennaro ML (2021) Highly versatile antibody binding assay for the detection of SARS-CoV-2 infection and vaccination. J Immunol Methods 499: 113165. PMI: 34634317
Ravi J, Anantharaman V, Zorn Chen S, Datta P, Aravind L, Gennaro ML (2020) Phage-shock-protein (Psp) Envelope Stress Response: Evolutionary History & Discovery of Novel Players. BioRxiv: preprint. PMI:
Guerrini V, Panettieri RA, Jr., Gennaro ML (2020) Lipid-laden macrophages as biomarkers of vaping-associated lung injury. Lancet Respir Med 8: e6. PMI: 32035068
Bruiners N, Dutta N, Guerrini V, Salamon H, Yamaguchi KD, Karakousis PC, Gennaro ML (2020) The anti-tubercular activity of simvastatin is mediated by cholesterol-driven autophagy via the AMPK-mTORC1-TFEB axis. J Lipid Res. PMI: 32848049
Guerrini V, Gennaro ML (2019) Foam Cells: One Size Doesn’t Fit All. Trends Immunol 40: 1163-1179. PMI: 31732284
Guerrini V, Prideaux B, Blanc L, Bruiners N, Arrigucci R, Singh S, Ho-Liang HP, Salamon H, Chen PY, Lakehal K, Subbian S, O’Brien P, Via LE, Barry CE, 3rd, Dartois V, Gennaro ML (2018) Storage lipid studies in tuberculosis reveal that foam cell biogenesis is disease-specific. PLoS Pathog 14: e1007223. PMI: 30161232
Arrigucci R, Lakehal K, Vir P, Handler D, Davidow AL, Herrera R, Estrada-Guzman JD, Bushkin Y, Tyagi S, Lardizabal AA, Gennaro ML (2018) Active Tuberculosis Is Characterized by Highly Differentiated Effector Memory Th1 Cells. Front Immunol 9: 2127. PMI: 30283456
Broger T, Basu Roy R, Filomena A, Greef CH, Rimmele S, Havumaki J, Danks D, Schneiderhan-Marra N, Gray CM, Singh M, Rosenkrands I, Andersen P, Husar GM, Joos TO, Gennaro ML, Lochhead MJ, Denkinger CM, Perkins MD (2017) Diagnostic Performance of Tuberculosis-Specific IgG Antibody Profiles in Patients with Presumptive Tuberculosis from Two Continents. Clin Infect Dis 64: 947-955. PMI: 28362937
Manganelli R, Gennaro ML (2016) Protecting from Envelope Stress: Variations on the Phage-Shock-Protein Theme. Trends Microbiol. PMI: 27865622
Dutta NK, Bruiners N, Pinn ML, Zimmerman MD, Prideaux B, Dartois V, Gennaro ML, Karakousis PC (2016) Statin adjunctive therapy shortens the duration of TB treatment in mice. J Antimicrob Chemother 71: 1570-1577. PMI: 26903278
Chauhan R, Ravi J, Datta P, Chen T, Schnappinger D, Bassler KE, Balazsi G, Gennaro ML (2016) Reconstruction and topological characterization of the sigma factor regulatory network of Mycobacterium tuberculosis. Nat Commun 7: 11062. PMI: 27029515
Ascensao JA, Datta P, Hancioglu B, Sontag E, Gennaro ML, Igoshin OA (2016) Non-monotonic Response to Monotonic Stimulus: Regulation of Glyoxylate Shunt Gene-Expression Dynamics in Mycobacterium tuberculosis. PLoS Comput Biol 12: e1004741. PMI: 26900694
Shi L, Salamon H, Eugenin EA, Pine R, Cooper A, Gennaro ML (2015) Infection with Mycobacterium tuberculosis induces the Warburg effect in mouse lungs. Sci Rep 5: 18176. PMI: 26658723
Salamon H, Bruiners N, Lakehal K, Shi L, Ravi J, Yamaguchi KD, Pine R, Gennaro ML (2014) Cutting edge: Vitamin D regulates lipid metabolism in Mycobacterium tuberculosis infection. J Immunol 193: 30-34. PMI: 24899504
Kunnath-Velayudhan S, Salamon H, Wang HY, Davidow AL, Molina DM, Huynh VT, Cirillo DM, Michel G, Talbot EA, Perkins MD, Felgner PL, Liang X, Gennaro ML (2010) Dynamic antibody responses to the Mycobacterium tuberculosis proteome. Proc Natl Acad Sci U S A 107: 14703-14708. PMI: 20668240
Shi L, Sohaskey CD, Kana BD, Dawes S, North RJ, Mizrahi V, Gennaro ML (2005) Changes in energy metabolism of Mycobacterium tuberculosis in mouse lung and under in vitro conditions affecting aerobic respiration. Proc Natl Acad Sci U S A 102: 15629-15634. PMI: 16227431
Shi L, Jung YJ, Tyagi S, Gennaro ML, North RJ (2003) Expression of Th1-mediated immunity in mouse lungs induces a Mycobacterium tuberculosis transcription pattern characteristic of nonreplicating persistence. Proc Natl Acad Sci U S A 100: 241-246. PMI: 12506197
Goulding CW, Parseghian A, Sawaya MR, Cascio D, Apostol MI, Gennaro ML, Eisenberg D (2002) Crystal structure of a major secreted protein of Mycobacterium tuberculosis-MPT63 at 1.5-A resolution. Protein Sci 11: 2887-2893. PMI: 12441386
Colangeli R, Spencer JS, Bifani P, Williams A, Lyashchenko K, Keen MA, Hill PJ, Belisle J, Gennaro ML (2000) MTSA-10, the product of the Rv3874 gene of Mycobacterium tuberculosis, elicits tuberculosis-specific, delayed-type hypersensitivity in guinea pigs. Infect Immun 68: 990-993. PMI: 10639479